ID:DDX3X_HUMAN DESCRIPTION: RecName: Full=ATP-dependent RNA helicase DDX3X; EC=3.6.4.13; AltName: Full=DEAD box protein 3, X-chromosomal; AltName: Full=DEAD box, X isoform; AltName: Full=Helicase-like protein 2; Short=HLP2; FUNCTION: Multifunctional ATP-dependent RNA helicase. The ATPase activity can be stimulated by various ribo- and deoxynucleic acids indicative for a relaxed substrate specificity. In vitro can unwind partially double stranded DNA with a preference for 5'- single stranded DNA overhangs. Is involved in several steps of gene expression, such as transcription, mRNA maturation, mRNA export and translation. However, the exact mechanisms are not known and some functions may be specific for a subset of mRNAs. Involved in transcriptional regulation. Can enhance transcription from the CDKN1A/WAF1 promoter in a SP1-dependent manner. Found associated with the E-cadherin promoter and can down-regulate transcription from the promoter. Involved in regulation of translation initiation. Proposed to be involved in positive regulation of translation such as of cyclin E1/CCNE1 mRNA and specifically of mRNAs containing complex secondary structures in their 5'UTRs; these functions seem to require RNA helicase activity. Specifically promotes translation of a subset of viral and cellular mRNAs carrying a 5'proximal stem-loop structure in their 5'UTRs and cooperates with the eIF4F complex. Proposed to act prior to 43S ribosomal scanning and to locally destabilize these RNA structures to allow recognition of the mRNA cap or loading onto the 40S subunit. After association with 40S ribosomal subunits seems to be involved in the functional assembly of 80S ribosomes; the function seems to cover translation of mRNAs with structured and non-structured 5'UTRs and is independent of RNA helicase activity. Also proposed to inhibit cap-dependent translation by competetive interaction with EIF4E which can block the EIF4E:EIF4G complex formation. Proposed to be involved in stress response and stress granule assembly; the function is independent of RNA helicase activity and seems to involve association with EIF4E. May be involved in nuclear export of specific mRNAs but not in bulk mRNA export via interactions with XPO1 and NXF1. Also associates with polyadenylated mRNAs independently of NXF1. Associates with spliced mRNAs in an exon junction complex (EJC)-dependent manner and seems not to be directly involved in splicing. May be involved in nuclear mRNA export by association with DDX5 and regulating its nuclear location. Involved in innate immune signaling promoting the production of type I interferon (IFN-alpha and IFN-beta); proposed to act as viral RNA sensor, signaling intermediate and transcriptional coactivator. Involved in TBK1 and IKBKE-dependent IRF3 activation leading to IFN-beta induction. Also found associated with IFN-beta promoters; the function is independent of IRF3. Can bind to viral RNAs and via association with MAVS/IPS1 and DDX58/RIG-I is thought to induce signaling in early stages of infection. Involved in regulation of apoptosis. May be required for activation of the intrinsic but inhibit activation of the extrinsic apoptotic pathway. Acts as an antiapoptotic protein through association with GSK3A/B and BIRC2 in an apoptosis antagonizing signaling complex; activation of death receptors promotes caspase-dependent cleavage of BIRC2 and DDX3X and relieves the inhibition. May be involved in mitotic chromosome segregation. Appears to be a prime target for viral manipulations. Hepatitis B virus (HBV) polymerase and possibly vaccinia virus (VACV) protein K7 inhibit IFN-beta induction probably by dissociating DDX3X from TBK1 or IKBKE. Is involved in hepatitis C virus (HCV) replication; the function may involve the association with HCV core protein. HCV core protein inhibits the IPS1- dependent function in viral RNA sensing and may switch the function from a INF-beta inducing to a HCV replication mode. Involved in HIV-1 replication. Acts as a cofactor for XPO1- mediated nuclear export of incompletely spliced HIV-1 Rev RNAs. CATALYTIC ACTIVITY: ATP + H(2)O = ADP + phosphate. SUBUNIT: Interacts with XPO1, TDRD3, PABPC1, NXF1, EIF3C, MAVS, DDX58 and NCAPH. Interacts with DDX5; the interaction is regulated by the phosphorylation status of both proteins. Interacts with EIF4E; DDX3X competes with EIF4G1/EIF4G3 for interaction with EIF4E. Interacts with IKBKE; the interaction is found to be induced upon virus infection and to be inhibited by HBV polymerase. Interacts with TBK1; the interaction is inhibited by HBV polymerase. Associates with the eukaryotic translation initiation factor 3 (eIF-3) complex. Associates with the 40S ribosome. Identified in a mRNP complex, at least composed of DHX9, DDX3X, ELAVL1, HNRNPU, IGF2BP1, ILF3, PABPC1, PCBP2, PTBP2, STAU1, STAU2, SYNCRIP and YBX1. Interacts with HCV core protein. Interacts with vaccinia virus (VACV) protein K7. Found in a complex with HIV-1 Rev and XPO1. INTERACTION: O95786:DDX58; NbExp=2; IntAct=EBI-353779, EBI-995350; P05198:EIF2S1; NbExp=3; IntAct=EBI-353779, EBI-1056162; P55884:EIF3B; NbExp=5; IntAct=EBI-353779, EBI-366696; Q99613:EIF3CL; NbExp=3; IntAct=EBI-353779, EBI-353741; Q7Z434:MAVS; NbExp=4; IntAct=EBI-353779, EBI-995373; Q15003:NCAPH; NbExp=2; IntAct=EBI-353779, EBI-1046410; Q9UBU9:NXF1; NbExp=5; IntAct=EBI-353779, EBI-398874; P11940:PABPC1; NbExp=9; IntAct=EBI-353779, EBI-81531; SUBCELLULAR LOCATION: Nucleus speckle. Cytoplasm. Mitochondrion outer membrane. Note=Located predominantly in nuclear speckles and, at low levels, throughout the cytoplasm. Located to the outer side of nuclear pore complexes (NPC). Shuttles between the nucleus and the cytoplasm in a XPO1 and may be also in a NFX1-dependent manner. Associated with polyadenylated mRNAs in the cytoplasm and the nucleus. Predominantly located in nucleus during G(0) phase and in the cytoplasm during G1/S phase. INDUCTION: Regulated by the cell cycle. Maximally expressed din the cytoplasm uring G1/S phase and decreased expression during G2/M phase. PTM: Phosphorylated by TBK1; the phosphorylation is required to synergize with TBK1 in IFN-beta induction. Probably also phosphorylated by IKBKE. The cytoplasmic form is highly phosphorylated in the G1/S phase and much lower phosphorylated in G2/M. SIMILARITY: Belongs to the DEAD box helicase family. DDX3/DED1 subfamily. SIMILARITY: Contains 1 helicase ATP-binding domain. SIMILARITY: Contains 1 helicase C-terminal domain.
The RNAfold program from the Vienna RNA Package is used to perform the secondary structure predictions and folding calculations. The estimated folding energy is in kcal/mol. The more negative the energy, the more secondary structure the RNA is likely to have.
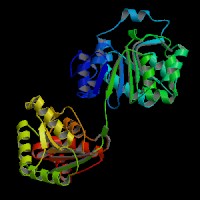
ModBase Predicted Comparative 3D Structure on O00571
Front
Top
Side
The pictures above may be empty if there is no ModBase structure for the protein. The ModBase structure frequently covers just a fragment of the protein. You may be asked to log onto ModBase the first time you click on the pictures. It is simplest after logging in to just click on the picture again to get to the specific info on that model.
Orthologous Genes in Other Species
Orthologies between human, mouse, and rat are computed by taking the best BLASTP hit, and filtering out non-syntenic hits. For more distant species reciprocal-best BLASTP hits are used. Note that the absence of an ortholog in the table below may reflect incomplete annotations in the other species rather than a true absence of the orthologous gene.
 Sequence and Links to Tools and Databases
Sequence and Links to Tools and Databases  Common Gene Haplotype Alleles
Common Gene Haplotype Alleles